
A3
Cat.No.: CSC-C9153W
Species: Homo sapiens (Human)
Source: Blood; Peripheral Blood
Morphology: lymphoblast
- Specification
- Background
- Scientific Data
- Q & A
- Customer Review
The A3 cell line is a human T-lymphoblast subclone derived from the well-established Jurkat acute T-cell leukemia line. It was generated by treating parental Jurkat cells with Fas antibody followed by limiting dilution cloning. A key biological feature of the A3 clone is its markedly heightened sensitivity to Fas (CD95/Apo-1)-mediated apoptosis compared with the parent line, and a very low spontaneous resistance rate, making it a refined model for death receptor signaling studies.
Morphologically, A3 cells appear as round lymphoblasts and grow in suspension. They express T-cell markers including CD3 and TCR complex components. The cells are routinely cultured in RPMI-1640 medium supplemented with serum and L-glutamine, under standard conditions (37°C, 5% CO₂), and are passaged by gentle pipetting and dilution (typically 1:2 to 1:4) when the density reaches ~0.5-1 × 10⁶ cells/mL.
A3 cells are primarily used to investigate T-cell receptor (TCR) signal transduction, Fas/CD95 apoptotic pathways, NF-κB and NFAT activation, and for in vitro screening of immunomodulatory or anti-leukemic compounds.
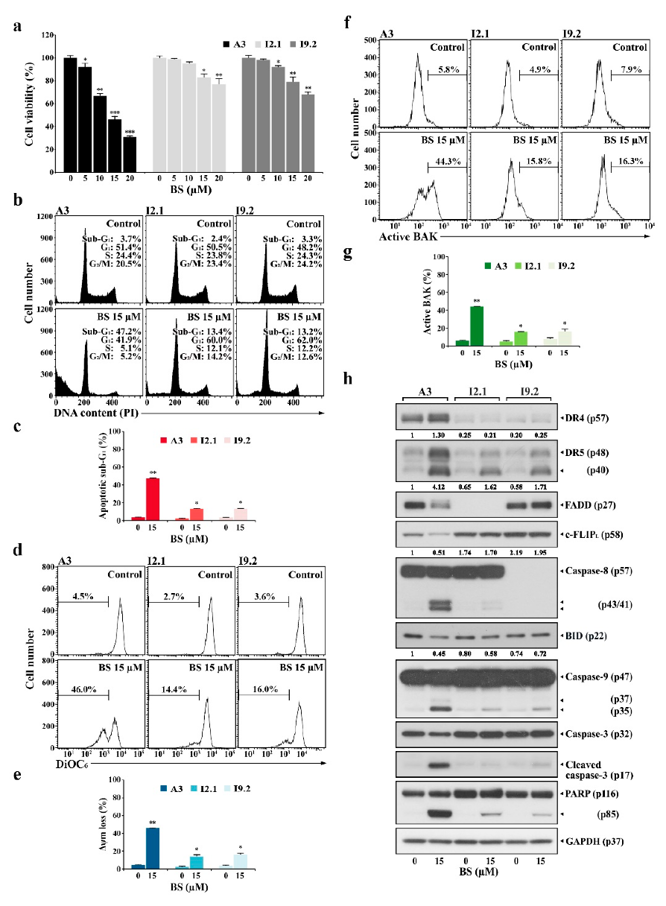
The CDK7 Inhibitor BS-181 Induces Mitochondrial Apoptosis Primarily via the Extrinsic Pathway in T-ALL
Chemotherapy resistance in T-cell acute lymphoblastic leukemia (T-ALL) results in poor prognosis. Park et al. elucidates the mechanism of the CDK7 inhibitor BS-181 using malignant cell lines (Jurkat A3, U937, HeLa) and normal human T cells.
To determine if BS-181 induces mitochondrial apoptosis strictly via the extrinsic pathway, they compared wild-type Jurkat A3 cells with mutants deficient in key extrinsic mediators: I2.1 (FADD-deficient) and I9.2 (caspase-8-deficient). Treatment with 5-20 µM BS-181 reduced A3 viability in a dose-dependent manner (91.7% to 30.7%), whereas I2.1 and I9.2 cells were significantly less affected (Fig. 1a).
Flow cytometry confirmed this differential sensitivity. At 15 µM BS-181, A3 cells exhibited a 47.2% sub-G1 population, 46.0% mitochondrial membrane potential (ΔΨm) loss, and 44.3% BAK activation (Fig. 2b-g). In contrast, I2.1 and I9.2 cells showed markedly lower apoptosis rates (sub-G1: 13.4% and 13.2%, respectively), reduced ΔΨm loss (14.4% and 16.0%), and diminished BAK activation (15.8% and 16.3%). Interestingly, while BS-181 did not cause G1 arrest in A3 cells, it induced cell cycle arrest in the resistant I2.1 and I9.2 mutants. These results demonstrate that BS-181 primarily triggers BAK-dependent mitochondrial apoptosis as a downstream consequence of extrinsic pathway activation. However, at higher concentrations (≥15 µM), BS-181 can also bypass the requirement for FADD and caspase-8 to induce apoptosis independently.
Ask a Question
Write your own review
- You May Also Need

Description: Established in 2007 from the bone marrow mononuclear cells of an 82-year-old Japanese man with diffuse large B-cell lymphoma in the leukemic phase

Description: Established from the bone marrow of a 28-year-old man who developed the terminal leukemic phase of lymphosarcoma in 1976
Description: This cell line was derived from the bone marrow aspirate of a 59 year old male with erythroleukemia that became acute myelogenous leukaemia.The cells form colonies in soft-agar in the presence of ...

Description: Established from the pleural effusion of a 24-year-old woman with recurrent anaplastic large cell lymphoma (ALCL); cells were described to clonally derive from T-lineage lymphoid cells and to be ...

Description: Established from a 37-year-old man at second (refractory/terminal) relapse of Hodgkin lymphoma (nodular sclerosing -> lymphocyte depleted/stage IIISA -> stage IV) after both combined chemo- and ...

Description: Established from the peripheral blood of a 10-year-old Caucasian boy with acute lymphoblastic leukemia (pre B-ALL) at diagnosis in 1993
- Adipose Tissue-Derived Stem Cells
- Human Neurons
- Mouse Probe
- Whole Chromosome Painting Probes
- Hepatic Cells
- Renal Cells
- In Vitro ADME Kits
- Tissue Microarray
- Tissue Blocks
- Tissue Sections
- FFPE Cell Pellet
- Probe
- Centromere Probes
- Telomere Probes
- Satellite Enumeration Probes
- Subtelomere Specific Probes
- Bacterial Probes
- ISH/FISH Probes
- Exosome Isolation Kit
- Human Adult Stem Cells
- Mouse Stem Cells
- iPSCs
- Mouse Embryonic Stem Cells
- iPSC Differentiation Kits
- Mesenchymal Stem Cells
- Immortalized Human Cells
- Immortalized Murine Cells
- Cell Immortalization Kit
- Adipose Cells
- Cardiac Cells
- Dermal Cells
- Epidermal Cells
- Peripheral Blood Mononuclear Cells
- Umbilical Cord Cells
- Monkey Primary Cells
- Mouse Primary Cells
- Breast Tumor Cells
- Colorectal Tumor Cells
- Esophageal Tumor Cells
- Lung Tumor Cells
- Leukemia/Lymphoma/Myeloma Cells
- Ovarian Tumor Cells
- Pancreatic Tumor Cells
- Mouse Tumor Cells